Session: (0066–0095) T Cell Biology & Targets in Autoimmune & Inflammatory Disease Poster
0084: Excess IL-18 Protects from Experimental Autoimmune Encephalomyelitis Mainly via Preferential Augmentation of Suppressor/Regulatory over Autoreactive T-cell Function
University of Pennsylvania, Perelman School of Medicine Philadelphia, PA, United States
Disclosure information not submitted.
Jeremy Morrissette1, Vinh Dang2, Jemy Varghese2, Zachary Lanzar1, Junior Nguyen2, Emily Landy3, Anastasia Frank-Kamenetskii4 and Scott Canna2, 1University of Pennsylvania, Perelman School of Medicine, Philadelphia, PA, 2Children's Hospital of Philadelphia, Philadelphia, PA, 3University of Pittsburgh, Pittsburgh, PA, 4CHOP/UPENN, Philadelphia, PA
Background/Purpose: Interleukin 18 (IL-18) is an inflammasome-activated cytokine that canonically amplifies interferon-gamma (IFNg) production and cytotoxicity by CD8 T-cells (CD8Ts) and Th1 CD4 T-cells (CD4Ts). It is inhibited by the circulating high affinity antagonist, IL-18 binding protein (Il18bp). In certain contexts, IL-18 amplifies non-Th1 responses including Th2, Th17, and Treg. Despite its likely pathogenic role in autoinflammation, clinical observations suggest a possible immunoregulatory role in CD4T-mediated autoimmunity. We sought to understand the role of excess IL-18 in the mixed Th1/17 experimental autoimmune encephalomyelitis (EAE) model of central nervous system autoimmunity.
Methods: EAE was induced via immunization with a complete Freund's adjuvant/Myelin Oligodendrocyte Glycoprotein peptide (MOG35-55) emulsion and pertussis toxin in the following mice: mice deficient in Il-18bp (Il18bp-/-), with transgenic overproduction of mature IL-18 (Il18tg), transgenic for a T-cell receptor recognizing MOG35-55 (2D2 mice), and relevant controls . EAE was monitored by daily clinical scoring of ascending paralysis. Cell number and cellular protein phenotype were determined by flow cytometry.
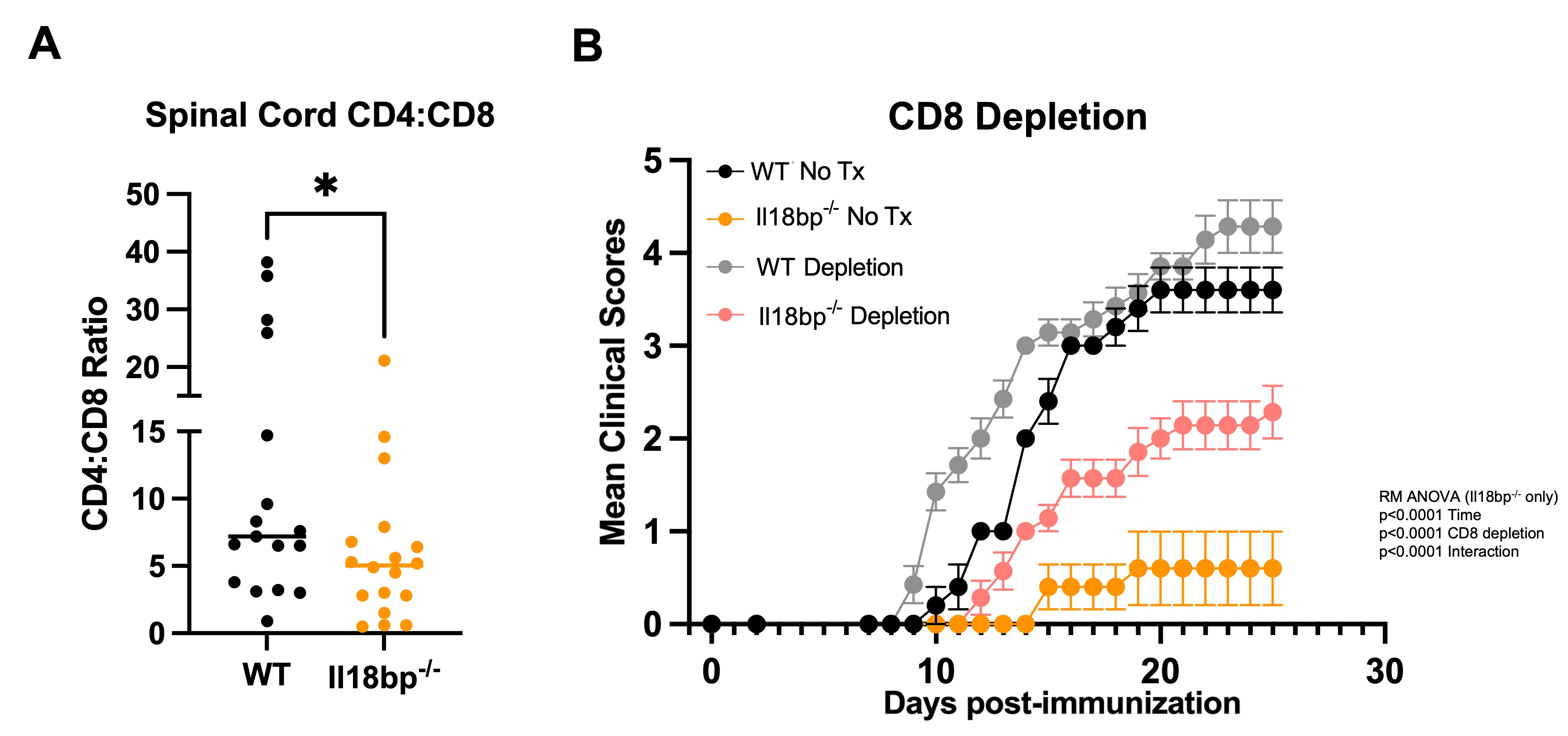
Results: We hypothesized that systemic excess IL-18, would amplify autoreactive T-cell activation and lead to more severe EAE, but both Il18bp-/- and Il18tg mice were profoundly protected from EAE (Fig. 1). Protection from EAE in Il18bp-/- mice depended on IFNg. Draining lymph nodes and spleens from Il18bp-/- mice showed a modest increase in IFNg-producing CD4Ts and equivalent IL-17-producing CD4Ts. Spinal cords demonstrated fewer CD4T cells but comparable markers of Th function/polarization on a per-cell basis. There was increased infiltration of CD8Ts relative to CD4Ts in spinal cords of Il18bp-/-mice (Fig. 2A). 2D2 mice have an abundance of MOG autoreactive CD4Ts and very few CD8Ts (due to allelic exclusion). Paradoxically, Il18bp-/-;2D2 mice developed more severe EAE than control 2D2 mice (Fig.1A), suggesting that an increase in precursor autoreactive CD4Ts and/or loss of CD8Ts switches the effect of excess IL-18 from protective to pathogenic. Sensitivity to EAE was restored in Il18bp-/-mice only after transfer of >5 million autoreactive 2D2 CD4Ts. Further, ex vivo culture with IL-18 improved CD8Ts ability to inhibit EAE while CD8-depletion substantially, but incompletely, diminished protection in Il18bp-/- mice (Fig.2B).
Conclusion: These data suggest the IL-18 can amplify both autoreactive CD4T and previously described CD8 T suppressor cells but the latter dominate with a physiologic T cell repertoire. Excess IL-18 may function to preferentially augment CD8T suppressor function to clear autoreactive CD4T and mediate protection in EAE. This may inform novel strategies to amplify endogenous suppressive T cells or enhance cellular therapeutics in the treatment of autoimmune diseases.
Fig. 1: Excess IL-18 protects against EAE development with a physiologic T cell repertoire. Il18bp-/-, 2D2, 2D2;Il18bp-/- (A), or Il18tg (B) mice underwent EAE immunization on day 0 with daily clinical EAE scoring compared to WT control mice.
Fig. 2: Excess IL-18 enriches CD8 T cells and depends on CD8 T cell function for protection. Spinal cords were analyzed by flow cytometry from WT and Il18bp-/- mice at peak disease (day 19 to 21) and the ratio of CD4 to CD8 T cells is represented (A). Mean clinical EAE score of WT and Il18bp-/- mice receiving isotype control (LIF2 Rat IgG2b) or CD8 depleting antibody (YTS169.4) q3d starting on day 0 (B).
J. Morrissette: None; V. Dang: None; J. Varghese: None; Z. Lanzar: None; J. Nguyen: None; E. Landy: None; A. Frank-Kamenetskii: None; S. Canna: Apollo Therapeutics, 2, Novartis, 12, Site PI for industry-sponsored trial, PracticePoint CME, 6, Simcha Therapeutics, 2, Sobi, 6.



.jpg)